





lncRNA新發現:PART1與ADAMTS9-AS2拮抗調控前列腺癌AR信號及細胞衰老機制
前列腺癌(PCa)最初是一種激素依賴性疾病,其發展和擴散與雄激素受體(AR)信號活性緊密相關。針對 AR 通路的治療,如使用 AR 拮抗劑和雙極雄激素療法中采用的超生理雄激素水平(SAL),是治療 PCa 的標準方法。在此,作者通過檢測 PCa 標本、觀察對 AR 拮抗劑或 SAL 的反應,以及進行功能缺失實驗,發現兩種長鏈非編碼 RNA(lncRNA)PART1 和 ADAMTS9-AS2 在一定程度上介導了 PCa 細胞中的雄激素信號傳導。結果顯示PART1 和 ERVH48-1 在 TT 樣本中顯著高表達,而 ADAMTS9-AS2 在 TT 樣本中的表達低于 NTAT 樣本。SAL 處理在兩種人類 PCa 細胞系和患者腫瘤樣本中呈現出相反的調控作用:ADAMTS9-AS2 表達上調,PART1 表達受抑制。此外,研究數據表明,ADAMTS9-AS2 可能作為 AR 信號的共激活因子,而 PART1 則作為共抑制因子。有趣的是,敲低實驗顯示,ADAMTS9-AS2 和 PART1 均能調節 AR 活性、蛋白水平,以及 SAL 介導的細胞衰老誘導過程。因此,數據表明 ADAMTS9-AS2 和 PART1 在 SAL 條件下對 AR 信號傳導起到調控作用。這篇文章于2025年3月發表于《International Journal of Surgery》期刊上,IF:12.5。
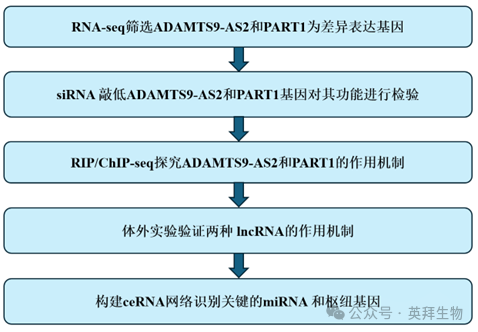
研究技術路線:
主要實驗結果:
1、患者前列腺癌樣本中的差異表達
PART1 在腫瘤組織(TT)與正常對照(NC)之間,以及 TT 與腫瘤旁正常組織(NTAT)之間的表達存在差異(圖 1)。ADAMTS9-AS2 在 NTAT 樣本中的表達高于 TT 和 NC 樣本。C1QTNF3-AMACR 在三組樣本中的表達沒有差異。C8orf34-AS1 在 NC 樣本中的表達高于其他組樣本。ERVH48-1在所有三組比較(TT/NC、TT/NTAT 和 NC/NTAT)中的表達均存在差異。最后,KIAA0087 和 ENO1-AS1 在 TT 與 NC 之間,以及 NC 與 NTAT 之間的表達水平不同(圖 1)。
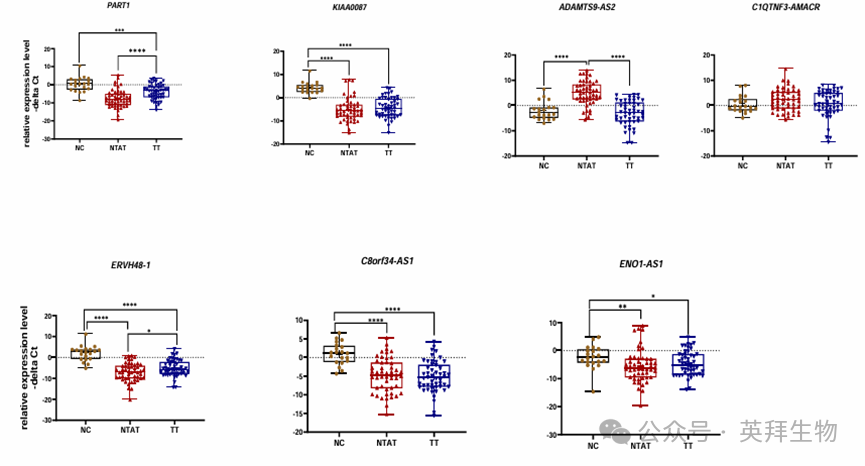
圖 1. 比較正常對照組織(NC)、腫瘤旁正常組織(NTAT)和前列腺腫瘤組織(TT)中所選 lncRNA 基因的不同表達模式
PART1 和 ERVH48-1 在 TT 樣本中的表達水平高于 NTAT 樣本(表達比值分別為13.1 和 3.6;P 值分別 < 0.0001 和 0.04)。值得注意的是,PART1、KIAA0087、ERVH48-1、C8orf34-AS1 和 ENO1-AS1 在 TT 樣本中的表達低于 NC 樣本(P 值分別為 0.0002、<0.0001、<0.0001、<0.0001 和 0.04)。雖然 PART1、KIAA0087、ERVH48-1、C8orf34-AS1 和 ENO1-AS1 在 NTAT 樣本中的表達低于 NC 樣本,但 ADAMTS9-AS2 在 NTAT 樣本中的表達高于 NC 樣本(表達比值(95% 置信區間) = 135(27.3 - 670),P 值 < 0.0001)。詳細統計數據見表 S4。這些數據表明,除C1QTNF3-AMACR 外,分析的基因在這些樣本中均表現出顯著不同的表達水平。基于 NTAT 和 NC 之間三個基因表達的顯著差異,數據還表明,NTAT 樣本衍生的數據不能總是被視為非腫瘤對照。
在 TT、NTAT 和 NC 樣本中,PART1、KIAA0087、ADAMTS9-AS2、C1QTNF3-AMACR、ERVH48-1、C8orf34-AS1 和 ENO1-AS1 的表達水平之間存在顯著的成對相關性。最強的相關性出現在 ENO1-ASI和 KIAA0087 之間(相關系數 = 0.95);以及 C8orf34-AS1 與另外兩個 lncRNA,即 KIAA0087 和 ERVH48-1 在 TT 樣本中的相關性(相關系數 = 0.95)。同樣,在這些樣本中,ENO1-ASI 的表達水平與 PART1 和 C8orf34-AS1 的表達水平相關,相關系數為 0.94。然后,作者使用層次聚類方法(Ward 法,歐氏距離)根據表達水平將lncRNA 分為四個簇。C1QTNF3-AMACR和 ADAMTS9-AS2 聚為一組;而 ERVH48-1 和 C8orf34-AS1 被識別為聚在另一組。最后,PART1 和 KIAA0087 構成了另一個簇。值得注意的是,大多數患者樣本(TT1-TT50 和 NTAT1-NTAT50)中分析的 lncRNA 表達水平降低。
受試者工作特征(ROC)曲線分析顯示,KIAA0087在區分 TT 與 NC 樣本(AUC = 0.96)以及 NTAT 與 NC 樣本(AUC = 0.93)方面表現出色。此外,ERVH48-1 在區分 TT 樣本與 NC 樣本(AUC = 0.91)以及 NTAT 樣本與 NC 樣本(AUC = 0.95)方面表現適宜。TT 樣本中 C1QTNF3-AMACR 的相對表達水平與體重指數(BMI)相關(P = 0.02)。此外,ADAMTS9-AS2 的表達水平與前列腺體積相關(P = 0.05)。前列腺癌患者的年齡與前列腺體積之間存在顯著正相關(x2=6.41),P = 0.01)。正如預期的那樣, Gleason 評分與前列腺特異性抗原(PSA)濃度之間也存在顯著正相關(=6.28,P = 0.04)。
2、SAL誘導ADAMTS9-AS2上調,而PART1下調
去勢抵抗性 C4-2 細胞的實驗顯示,用 SAL 處理后,ADAMTS9-AS2 顯著上調。用第二代 AR 拮抗劑恩雜魯胺(ENZ)和達羅他胺(ODM)處理對該 lncRNA 的表達沒有顯著影響(圖 2)。另一方面,用 SAL 處理后,PART1 的表達下調,用 AR 拮抗劑 ENZ 或 ODM 處理也會使其下調。通過分析作者之前的 RNA 測序數據,證實了 SAL 對 PART1 的下調作用。
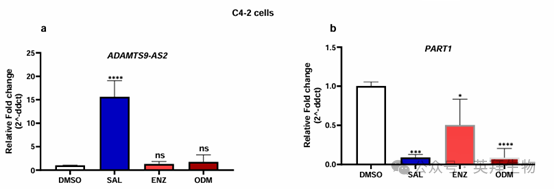
圖 2. SAL 處理增強了 lncRNA ADAMTS9-AS2 的表達,而 lncRNA PART1 的表達則被 SAL 和第二代 AR 拮抗劑抑制
這表明這兩個 lncRNA 是 AR 信號通路的一部分,也表明AR 拮抗劑并不總是表現出與雄激素相反的調節作用,并且對于 PART1 而言,AR 拮抗劑可能不會使 AR 失活,反而可能賦予 AR 轉錄活性。
在雄激素敏感的 LNCaP 細胞中進行的類似實驗顯示,用 SAL 處理可有效誘導ADAMTS9-AS2 的表達。用 Enz 或 ODM 處理也觀察到表達上調(圖 3)。相反,用 SAL 或 AR 拮抗劑處理后,LNCaP 細胞中 PART1 的表達下調。因此,數據表明 PART1 和 ADAMTS9-AS2 的表達處于 AR 信號通路中。有趣的是,PART1 的表達被 SAL 或 AR 拮抗劑抑制,而 ADAMTS9-AS2 的表達則被 AR 配體(包括 AR 拮抗劑)激活。
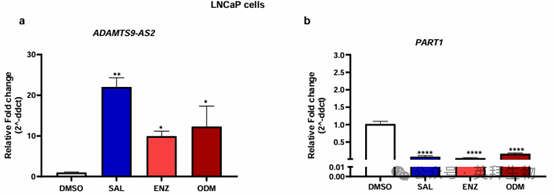
圖 3. SAL 和 AR 拮抗劑增強了 lncRNA ADAMTS9-AS2 的表達,而 lncRNA PART1 的表達則被 SAL 和 AR 拮抗劑抑制
3、3D腫瘤球體模型揭示AR調節治療的差異效應
與傳統的二維貼壁培養相比,3D 腫瘤球體模型被認為能更準確地反映腫瘤的復雜性。為了更好地模擬腫瘤微環境,作者使用超低成本貼壁板,從親本 C4-2 或 LNCaP 細胞生成 LNCaP 和 C4-2 細胞的 3D 腫瘤球體模型。該模型提供了一個更符合生理相關性的系統,用于評估雄激素受體(AR)調節治療(包括 SAL、ENZ 和 ODM)相對于 DMSO 對照的影響。
對球體體積的定量評估顯示,用 SAL、ENZ 或 ODM 處理后,LNCaP 和 C4-2 球體的腫瘤大小均顯著減小(圖 4A,4B)。這一減少與之前在二維培養系統中的觀察結果一致,強調了這些藥物在破壞腫瘤生長方面的有效性。
對關鍵 AR 調節基因和 lncRNA 的分子分析顯示出治療特異性的表達模式。ADAMTS9-AS2,一種先前被認為參與 AR 信號調節的 lncRNA,在用 SAL、ENZ 和 ODM 處理后,在 LNCaP 和 C4-2 球體中的表達均顯著增加(圖 4A,4B)。這種模式支持其作為 AR 通路中潛在共調節因子的作用,并表明 AR 拮抗劑不會完全使 AR 信號失活。
相反,AR 靶基因,包括 TMPRSS2、KLK3 和 FKBP5 的表達,表現出對治療類型的明顯依賴性。這些 AR 靶基因在用 SAL 處理后上調,但在用 ENZ 和 ODM 處理的兩種細胞系中均下調(分別見圖 4A 和 4B)。這些差異反應突出了 SAL 作為 AR 激動劑與 ENZ/ODM 作為 AR 拮抗劑的對比效應。
有趣的是,lncRNA PART1 在用 SAL、ENZ 或 ODM 處理的 LNCaP 和 C4-2 球體中均顯示出持續下調(圖4A,4B)。這一發現與其作為 AR 信號共抑制因子的作用一致,特別是在 AR 通路調節的條件下。
這些結果共同突出了 3D 球體模型捕捉腫瘤特異性基因表達對 AR 靶向治療反應復雜性的能力。它們還進一步證明了 AR 信號與 lncRNA 調節之間的相互作用,強調了它們在前列腺癌治療中的相關性。
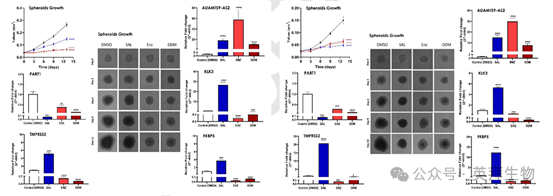
圖 4A 和圖 4B. SAL、ENZ 和 ODM 處理抑制 C4-2 和 LNCaP 細胞中的球體生長,并調節腫瘤球體中 ADAMTS9-AS2 和 PART1 的表達
4、C4-2細胞中AR對PART1和ADAMTS9-AS2基因座的差異招募
染色質免疫沉淀測序(ChIP-seq)分析表明,在 C4-2 細胞中,雙氫睪酮(DHT)可使 AR 招募到 PART1(內含子)基因座(圖 S4A)。此外,ChIP-seq 結果顯示,在經 DHT 處理的 C4-2 細胞中,AR 選擇性地招募到 ADAMTS9-AS2(內含子和外顯子)基因座。因此,數據表明 PART1 是一種新型的 AR 靶基因,然而,它是一種非經典的 AR 靶基因,其表達被 SAL 處理抑制。
5、體外分析揭示SAL誘導ADAMTS9-AS2和PART1表達的顯著變化
對前列腺切除術后未固定的原生患者前列腺癌標本進行 SAL 處理后的表達變化分析。作者檢測到用 SAL 處理后,ADAMTS9-AS2和 PART1 的體外表達水平存在顯著差異(圖 5),證實了 SAL 對 LNCaP 和 C4-2 細胞中這些 lncRNA 的調節作用。
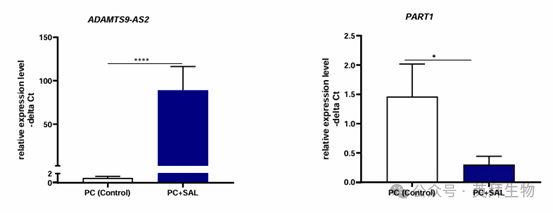
圖 5. 在原生人類前列腺癌組織中,SAL 處理可控制ADAMTS9-AS2 和 PART1 lncRNA的表達
與 SAL 誘導 ADAMTS - AS2 表達類似,在經 SAL 處理的前列腺癌組織中,ADAMTS9-AS2 的體外表達顯著升高(表達比值(95% 置信區間) = 54.2(16.9 - 175),P < 0.0001)。與 ADAMTS - AS2 相反,正如細胞系數據所預期的那樣,經 SAL 處理的前列腺癌組織中 PART1 的體外表達降低(表達比值(95% 置信區間) = 0.12(0.02 - 0.77),P = 0.03)。此外,還檢測到 PSA 水平與 BMI 之間存在顯著相關性(相關系數 = 0.726,P = 0.027),PART1 表達與年齡之間存在顯著相關性(相關系數 = -0.644,P = 0.049),以及 PART1 和 ADAMTS9-AS2 的表達水平之間存在顯著相關性(相關系數 = 0.8,P = 0.10)。
6、lncRNA ADAMTS9-AS2是AR的共調節因子
獲得的數據提出了一個假設,即 lncRNA ADAMTS9-AS2 與 AR 之間可能存在相互作用,影響AR 的轉錄活性。為此,通過小干擾RNA(siRNA)分別敲低 lncRNA ADAMTS9-AS2 和 PART1,并分析編碼診斷標志物前列腺特異性抗原(PSA)的 KLK3、TMPRSS2 和 FKBP5(已知的直接 AR 靶基因)的表達。在兩種細胞系中均驗證了敲低效果(圖 6a,6b),表明敲低有效。如預期的那樣,AR 靶基因(FKBP5(b)、TMPRSS2(c)和 KLK3(d))被 SAL 上調。有趣的是,ADAMTS9-AS2的敲低降低了 SAL 誘導的 AR 靶基因 FKB5、TMPRSS 和 KLK3 的表達。這一觀察結果表明,lncRNA ADAMTS9-AS2 可能作為 AR 的共激活因子發揮作用。因此,數據表明 lncRNA ADAMTS9-AS2 是 AR 信號通路的一部分。
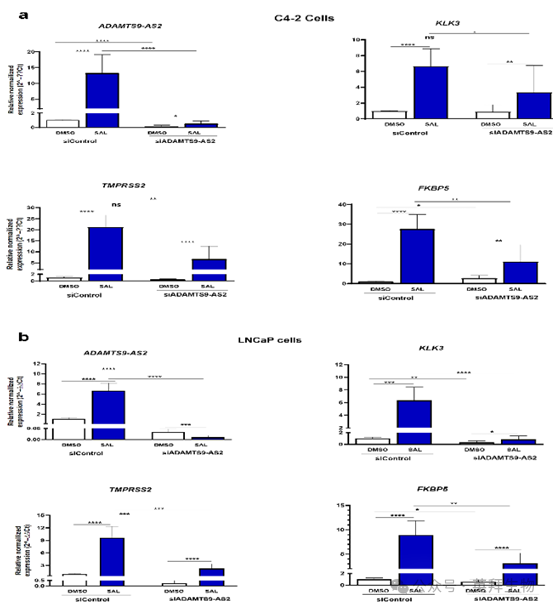
圖 6a 和圖 6b. ADAMTS9-AS2 的敲低降低了 AR 靶基因的表達
7、PART1 lncRNA敲低作為共抑制因子引發AR靶基因的過表達
同樣,作者在 C4-2 和 LNCaP 細胞系中對 PART1 進行了敲低。與 ADAMTS-AS2 的敲低情況相反,長鏈非編碼 RNA PART1 的表達受 SAL 抑制。敲低長鏈非編碼 RNA PART1 會導致 AR 靶基因表達增強(圖 7),這表明 PART1 是一種共抑制因子。與該觀察結果一致的是,在患者前列腺腺癌數據集(GEPIA)中,PART1 的表達與 KLK3和 FKBP5呈顯著正相關,與 TMPRSS2 的相關性較弱但也顯著。因此,這些數據表明,長鏈非編碼 RNA PART1可能作為一種共調節因子參與調節 AR12。
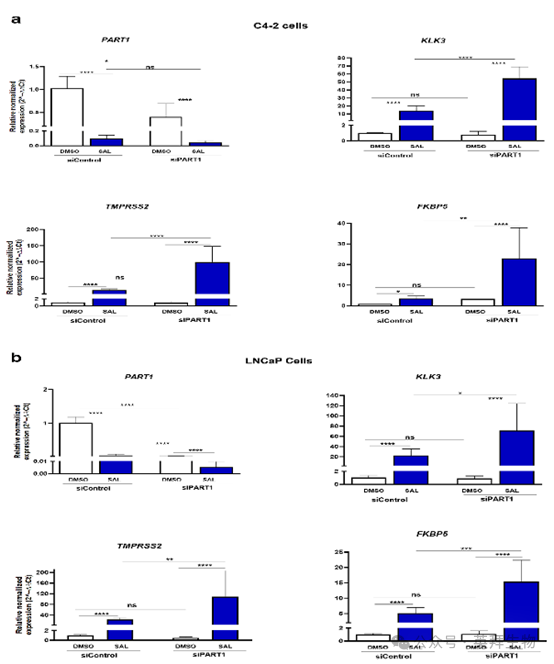
圖 7a 和圖 7b. PART1 的敲低
8、C4-2細胞中PART1和ADAMTS9-AS2的相互調節:對AR 信號傳導和細胞衰老的影響
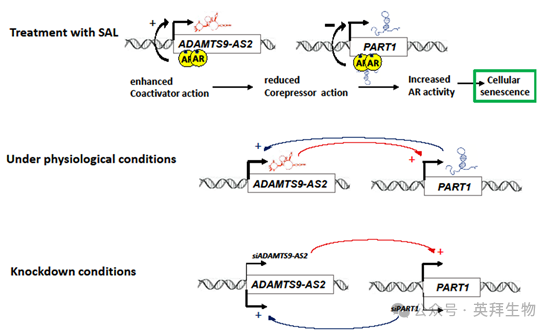
在通過 siRNA 介導敲低后,對 C4-2 細胞中 PART1 和 ADAMTS9-AS2 的表達分析揭示了一種動態調節的相互作用,這種相互作用影響著 AR 信號傳導(圖 8)。在沒有 SAL 的情況下,siADAMTS9-AS2 敲低會導致 PART1 表達增加,PART1 作為共抑制因子抑制 AR 靶基因。相反,siPART1 敲低會增強 ADAMTS9-AS2 的表達,促進 AR 激活。這表明這兩種長鏈非編碼 RNA 之間存在相互作用。在 SAL 處理后,ADAMTS9-AS2 水平上升而 PART1 水平下降,進一步證實了一種相互的交互作用,即一種長鏈非編碼 RNA 的增加對應著另一種的抑制。這種相互作用表明 AR 被 “超級激活”,從而驅動細胞衰老。
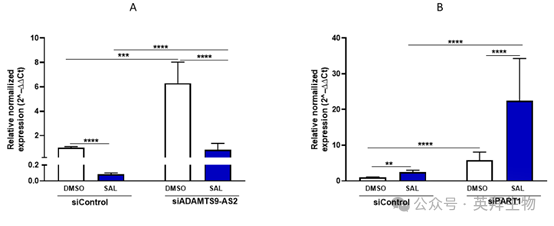
圖 8 C4-2 細胞中經 siRNA 介導敲低和 SAL 處理后 PART1 和 ADAMTS9-AS2 長鏈非編碼 RNA 的差異表達
9、RIP 測定突出了不同的富集模式:PART1被AR抗體顯著富集,而ADAMTS9-AS2則沒有
作者的結果揭示了在 LNCaP 細胞中使用 AR 抗體進行免疫沉淀時,沉淀物中存在獨特的富集模式。值得注意的是,PART1 轉錄本在使用抗 AR 抗體免疫沉淀的物質中豐度大幅增加,這表明它與 AR 存在相互作用。與之形成鮮明對比的是,ADAMTS9-AS2 沒有顯示出顯著的富集,這表明它與 AR 缺乏直接相互作用(圖 9)。
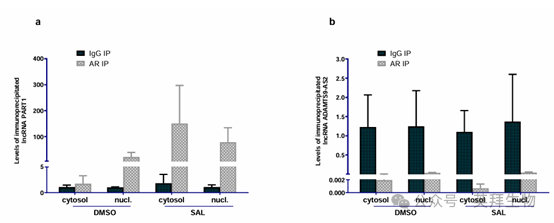
圖 9. AR 與長鏈非編碼 RNA PART1 結合。使用抗 AR 抗體通過 RIP 分析檢測 RNA 結合蛋白(RBP)AR 與 PART1 和 ADAMTS9-AS2 長鏈非編碼 RNA 的關聯
10、siRNA介導的ADAMTS9-AS2敲低降低了SAL誘導細胞衰老的能力
用 SAL 處理前列腺癌細胞會誘導細胞衰老,這在體外處理的患者前列腺癌標本、前列腺癌 3D 腫瘤球體以及小鼠異種移植物中均有體現。由于長鏈非編碼 RNA 調節 AR 信號傳導,作者假設它們調節 SAL 介導的細胞衰老誘導過程。通過 siRNA 介導的敲低實驗,作者的結果顯示,靶向降低 ADAMTS9-AS2 的表達會顯著減弱 SAL 在兩種細胞系中誘導細胞衰老的能力(圖 10)。這表明,被 SAL 上調的 ADAMTS9AS2 促進了 SAL 介導的細胞衰老。在研究 siRNA 介導的 PART1 敲低的影響時,作者觀察到兩種細胞系對 SAL 誘導的細胞衰老水平的細胞反應均有所降低。具體而言,在 C4-2 和 LNCaP 細胞中,降低 PART1 的表達會顯著降低 SAL 誘導細胞衰老的能力。這種增強的敏感性意味著 PART1 和 ADAMTS9-AS2 在雄激素敏感和去勢抵抗的前列腺癌細胞系中,在 AR 介導的衰老過程中均發揮著調節作用。
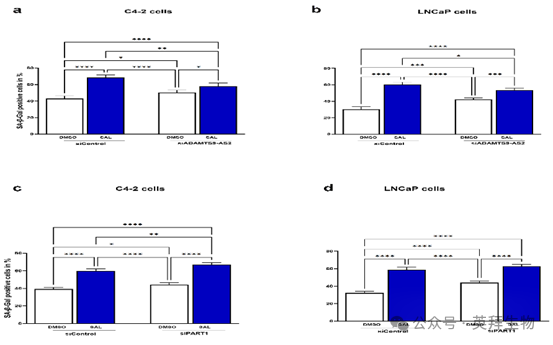
圖 10. 敲低長鏈非編碼 RNA ADAMTS9-AS2 會降低,而敲低 PART1 會增強 SAL 介導的細胞衰老
11、ADAMTS9-AS2和PART1敲低對C4-2 和LNCaP細胞中p15INK4B mRNA和蛋白水平的不同影響
在 C4-2 和 LNCaP 細胞中,分析了編碼 p15INK4B 蛋白的 CDKN2B mRNA 水平,實驗分為敲低 ADAMTS9-AS2 和未敲低的情況。結果表明,SAL 上調的 p15INK4B 的 mRNA 和蛋白水平均有所降低(圖 11A - C),并且敲低 ADAMTS - AS2 會下調兩種細胞系中 CDKN2B mRNA 和 p15INK4B 蛋白的水平(圖 11C)。這一發現表明 ADAMTS9-AS2 在衰老誘導因子 p15INK4B 的表達中發揮調節作用。作者的數據也與衰老相關 β- 半乳糖苷酶(SA-β-gal)檢測結果一致,該檢測也顯示敲低 ADAMTS9-AS2 后細胞衰老減少。因此,作者的觀察結果進一步證實,敲低ADAMTS9-AS2 會降低 SAL 誘導衰老的能力,并表明 ADAMTS9-AS2 在一定程度上介導了 SAL 誘導的細胞衰老。
與 si - ADAMTS9-AS2 的情況相反,在 LNCaP 和 C4-2 前列腺癌細胞系中敲低PART1,會顯著誘導p15INK4B 的 mRNA 和蛋白水平升高。這一觀察結果強調了 PART1 在調節 p15INK4B 表達中的作用,表明 PART1 通常可能作為 p15INK4B 表達的抑制因子。作者還分析了另一種細胞周期抑制劑 p21,結果表明,與 ADAMTS9-AS2 不同,敲低長鏈非編碼 RNA PART1 會增強 p21 蛋白水平,這表明 p21 也是 PART1 的作用靶點。有趣的是,在存在 SAL 的情況下,由于敲低兩種長鏈非編碼 RNA 會導致細胞衰老減少,p21 水平也會降低(圖 11C)。
這些發現表明,SAL 介導的 PART1 下調是前列腺癌細胞中 SAL 介導的細胞衰老誘導過程的一部分。敲低 PART1 后 p15NK4B 的上調也表明了 PART1 促進前列腺癌進展,甚至可能增強其侵襲性的潛在機制。與該觀察結果一致,來自患者前列腺癌樣本的數據集顯示 PART1 與 CDKN2B 之間存在顯著的負相關。值得注意的是,在兩種細胞系中,敲低 PART1 或 ADAMTS9-AS2 都會增強 AR 的蛋白水平(圖 11C),這表明這兩種長鏈非編碼 RNA 可能通過調節內源性 AR 蛋白水平來調節 AR 活性16。
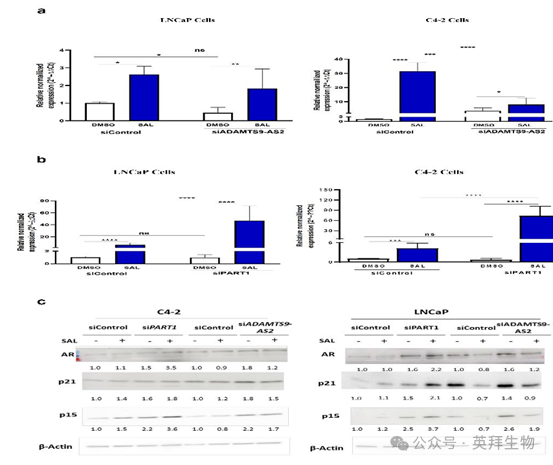
圖 11. 細胞周期抑制劑和衰老標志物 p15 的水平受 SAL 以及敲低長鏈非編碼 RNAs ADAMTS9-AS2 和 PART1 的影響
12、前列腺癌中ceRNA 網絡的構建
在構建包含長鏈非編碼 RNA(lncRNA)、微小 RNA(miRNA)和信使 RNA(mRNA)的競爭性內源性 RNA(ceRNA)網絡時,研究人員聚焦于表達上調或下調的 miRNA。如果在 miRNA - mRNA 和 lncRNA - miRNA 相互作用對中,lncRNA 和 mRNA 與這些 miRNA 呈現相反的關聯關系,那么它們就會被納入研究范圍。作者運用 miRcode來探究 lncRNA 和 miRNA 之間的聯系。分析結果顯示,在 9 種差異表達的 lncRNA 中,有 6 種可能靶向 55 種 miRNA 中的 17 種。作者使用 miRNAmeConverter 軟件包(版本 1.30.0),將 miRNA 的名稱轉換為 miRbase v22 數據庫中對應的名稱。隨后,利用 multimiR 軟件包(版本 1.24.0)來識別 miRNA 的靶 mRNA。為了優化結果,作者剔除了那些不在差異表達 mRNA 范圍內的 miRNA 靶向 mRNA。通過數據分析,作者確定了 13 種與前列腺癌相關的 miRNA,它們有可能靶向 266 種差異表達 mRNA 中的 107 種。此外,依據 ceRNA 網絡理論,作者去除了 lncRNA 與 miRNA 之間、miRNA 與 mRNA 之間存在相反表達模式的相互作用。借助 Cytoscape 3.9 軟件,作者構建了一個涵蓋 3 種 lncRNA、13 種 miRNA 和 107 種 mRNA 的綜合 ceRNA 網絡。該網絡包含 123 個節點和 124 條邊。
作者使用 cytoHubba插件,基于最大團中心性、度中心性、介數中心性、徑向中心性和離心率中心性,確定了 15 個樞紐基因。進一步分析這些樞紐基因的交集后,作者發現了一組包含 ADAMTS9 - AS2、hsa - miR - 10b - 5p、hsa - miR - 148a - 3p、hsa - miR - 183 - 5p、hsa - miR - 18b - 5p、hsa - miR - 199b - 5p、hsa - miR - 200a - 3p、hsa - miR - 203a - 3p、hsa - miR - 26b - 5p、hsa - miR - 301a - 3p、hsa - miR - 454 - 3p、hsa - miR - 7 - 5p 和 hsa - miR - 98 - 5p 的基因。作者還根據 ceRNA 網絡中的 mRNA 構建了蛋白質 - 蛋白質相互作用(PPI)網絡,并對該網絡應用中心性分析。最后,通過共享這些中心性樞紐基因之間的相互作用,作者得到了 CAV1 和 FLNA 這兩個樞紐 mRNA(圖 11)。這些樞紐基因在網絡中起著關鍵作用,對于理解它們在研究體系中的重要性至關重要。
對前列腺腺癌數據集的分析表明,CAV1 和 FLNA 的高表達水平與患者更高的生存概率呈正相關。這一發現凸顯了這兩個樞紐基因在前列腺癌病理生理學中的潛在重要性。該相關性與作者的生物信息學分析結果高度吻合,后者將 CAV1 和 FLNA 確定為調控網絡的關鍵組成部分。這些結果表明,這些基因的上調可能通過調節參與腫瘤進展、細胞結構或轉移行為的關鍵分子通路,為患者帶來生存優勢。不過,還需要進一步的實驗驗證,來闡明 CAV1 和 FLNA 的確切機制作用,并探索它們作為前列腺癌管理中的預后生物標志物或治療靶點的潛力。
對前列腺腺癌患者中 CAV1和 FLNA 表達進行的 Kaplan - Meier 生存分析顯示,它們與患者的預后存在顯著關聯。高 CAV1 表達(n = 334)與較低表達(n = 146)相比,患者的生存概率更高,這表明它有可能成為一種良好的預后標志物。同樣,FLNA 的高表達與更好的總體生存率相關,這表明它在前列腺癌進展過程中具有保護作用。這些發現突出了 CAV1 和 FLNA 在前列腺腺癌預后方面的重要性。
13、ceRNA網絡的關鍵軸
為了選擇關鍵軸,作者通過樞紐基因之間的交集來獲取基因。具體來說,在 ceRNA 網絡中,作者利用樞紐基因之間的共享相互作用來選擇 lncRNA 和 miRNA;對于 mRNA 的選擇,作者則利用 ceRNA 網絡中 mRNA 的 PPI 網絡里基因樞紐之間的共享相互作用。包括 CAV1 和 FLNA 在內的 mRNA 位于主要 PPI 網絡的最終基因樞紐中。這些關鍵軸包括 ADAMTS9 - AS2/hsa - miR - 7 - 5p/CAV1 和 ADAMTS9 - AS2/hsa - miR - 98 - 5p/FLNA(圖 12)。
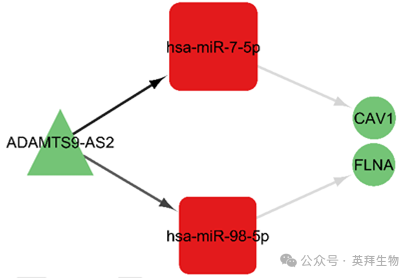
圖 12. 基于樞紐基因的 ceRNA 網絡關鍵軸
14、lncRNA - miRNA相互作用
此外,作者利用LncBase 數據庫,探究了 miRNA 與 ADAMTS9 - AS2 和 PART1 之間的相互作用。作者實施了多種篩選條件來優化分析,例如通過組織篩選進行聚焦,將 miRNA 置信度篩選設置為 “高” 以確保 miRNA 相互作用的可靠性,并使用 “已驗證” 篩選條件,僅納入經過實驗驗證的相互作用。分析結果顯示,PART1 與兩種 miRNA,即 hsa - miR - 99a - 5p 和 hsa - miR - 150 - 5p 之間存在顯著相互作用。此外,作者還發現 ADAMTS9 - AS2 與兩種 miRNA,hsa - miR - 7 - 5p 和 hsa - miR - 98 - 5p 之間存在值得關注的相互作用。
15、lncRNA相互作用
通過 RNAinter 數據庫,作者識別出 ADAMTS9 - AS2 和 PART1 與組蛋白修飾、轉錄因子(TFs)、蛋白質和 miRNA 的多種相互作用,這表明它們在染色質動態變化和轉錄調控中發揮著調節作用。ADAMTS9 - AS2 與活躍染色質標記,如H3K4me3(得分:0.8377)和 H3K27ac(0.7737),以及抑制性標記 H3K27me3(0.7579)都有很強的關聯,這表明它在激活和抑制基因表達方面具有雙重作用。它還與關鍵轉錄因子,包括 CTCF(0.6255)和 AR(0.4822),以及蛋白質 POLR2A(0.528)相互作用,這凸顯了它作為染色質和轉錄調節因子的潛力。同樣,PART1 與活躍組蛋白標記H3K4me3(0.6952)相關,并與轉錄因子,如 FOXA1(0.5984)和 AR(0.5479)相互作用,這表明它參與了轉錄調控,尤其是在激素驅動的通路中。這些發現突出了 lncRNA 在調節前列腺癌基因表達和細胞過程中的關鍵作用。
16、miR - 98 - 5p的表達上調,與ADAMTS9 - AS2存在顯著相關性
為了驗證生物信息學分析的結果,作者檢測了臨床樣本中 miR - 98 - 5p 和 miR - 7 - 5p 的表達水平(圖 13A)。與 NTAT 樣本相比,這兩種 miRNA 在 TT 樣本中的表達均下調(表達比值(95% 置信區間)分別為0.033±0.76(0.011 - 0.097)和 29.65±0.6(12.85 - 68.4);miR - 98 - 5p 和 miR - 7 - 5p 的 P 值分別 < 0.0001 和 0.001)。另一方面,與 NTAT 樣本相比,miR - 99a - 3p 和 miR - 150 - 5p 在 TT 樣本中的表達也下調(表達比值(95% 置信區間)分別為19.22±0.46(10.1 - 36.8)和 0.32±0.56(0.15 - 0.71);miR - 99a - 3p 和 miR - 150 - 5p 的 P 值分別 < 0.0001 和 0.006)。生物信息學分析與患者數據集驗證的結合,揭示了 ADAMTS9AS2 的一條新通路和一個新的 miRNA 網絡。
為了進一步探究潛在的相互作用,作者進行了 Spearman 相關性分析。在前列腺癌患者的腫瘤組織樣本中,miR - 99a - 3p 和 miR - 150 - 5p 的表達水平與 PART1 的相關性較弱(圖 13B)。同樣,miR - 7 - 5p 和 miR - 99 - 5p 與 ADAMTS9 - AS2 的相關性也較弱(圖 13D)。盡管這些相關性未達到統計學顯著性,但觀察到的趨勢,結合生物信息學預測和表達驗證,表明存在一個涉及 ADAMTS9 - AS2 的潛在 miRNA 網絡,值得進一步研究。
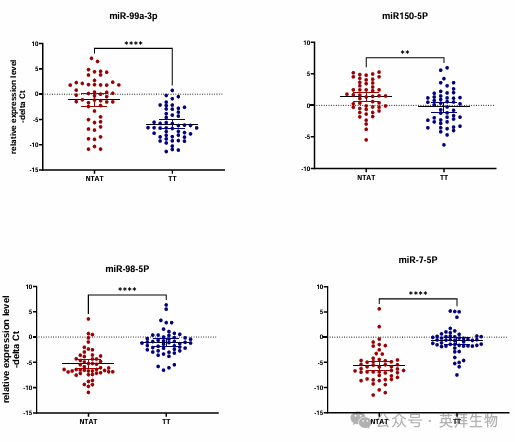 圖 13A. 使用患者樣本驗證生物信息學預測結果
圖 13A. 使用患者樣本驗證生物信息學預測結果
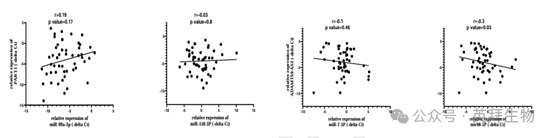
圖 13B. 前列腺癌樣本中 miRNA 與 lncRNA 的相關性分析
結論:
綜上所述,作者發現了新的 AR 信號通路,這些通路涉及在前列腺癌腫瘤發生過程中被 SAL 反向調節的長鏈非編碼 RNA。在雙極雄激素治療中使用的SAL,可誘導長鏈非編碼 RNA ADAMTS9 - AS2 的表達,同時抑制長鏈非編碼 RNA PART1 的表達。這兩種長鏈非編碼 RNA 均在 AR 信號通路中發揮作用,調控 SAL 介導的細胞衰老誘導過程。盡管它們受 SAL 的調控作用相反,但都參與了AR 信號傳導,這表明在前列腺癌的AR 信號通路中存在新的途徑和網絡。這些發現有助于理解BAT所涉及的分子機制,為前列腺癌的治療提供了潛在的新靶點和理論依據。不過,本研究存在一定的局限性,未來研究應注重體內驗證,并采用患者來源的異種移植或類器官模型,以便更深入地探究這些發現的臨床相關性。
實驗方法:
差異表達分析、GO分析、通路富集分析、構建競爭性內源RNA(ceRNA)網絡、相關性分析、生存分析、ChIP-seq、q-PCR、Western blot、RIP、細胞培養,siRNA敲低細胞轉染、3D腫瘤球體模型構建、細胞衰老誘導實驗、小鼠異種移植物實驗
參考文獻:
Taheri M, Schindler K, Baniahmad A. The LncRNAs PART1 and ADAMTS9-AS2 act in an Antithetic Manner on AR Signaling and Induction of Cellular Senescence in Prostate Cancer Cells. Int J Surg. 2025 Mar 26. doi: 10.1097/JS9.0000000000002334. Epub ahead of print. PMID: 40143747.